
So we come to the home stretch. Wulatelong gobiensis was named by Xu Xing, Lü Jun-chang and colleagues, and incorporates a peculiar array of caenagnathid and oviraptorid characters. My previous perspectives focused on the foot (preserved hinting at a hyperextensible second toe), the oddly caenagnathid-like hand, and the skull, which when correctly sized shows a small-headed animal. The authors argue that the holotype is a distinct, but basal, oviraptorid. This is based on their argument that several features of the skull and postcrania seem to occur in basal oviraptorosaurs, but not really in most oviraptorids. This post will now attempt to evaluate these qualities, and clarify if the components of cranial, appendicular/girdle and axial can clarify phylogenetic relationships (for example, whether this specimen is perhaps a basal oviraptorid retaining caenagnathoid-symplesiomorphies altered in caenagnathids, a basal caenagnathid retaining caenagnathoid-symplesiomorphies altered in oviraptorids, or perhaps even a descendant of the sister group of Caenagnathoidea. I have performed no new cladistic analysis taking these things into account.

“Shrunken head” version of Wulatelong gobiensis, with skull shown at preserved size. This is probably what Wulatelong looked like, but in the previous post I argued otherwise.
These features are:
1. external narial fenestra (ENF) less dorsally-positioned;
2. orbit rostrocaudally longer than infratemporal fenestra (ITF);
3. dorsal border of ITF narrower than ventral border;
4. anterior and posterior processes of lacrimal “intermediate” between basal oviraptorosaurs and oviraptorids;
5. ectopterygoid positioned rostrally;
6. external mandibular fenestra (EMF) is caudally positioned;
7. scapula shorter and more slender;
8. pubic peduncle of ilium wider than ischiadic peduncle;
9. ischium relatively short;
10. metatarsal III “high reduced,” by which it is meant MTIII is narrower in its dorsal extent, “pinched” between MTs II and IV.
Xu et al. further diagnose Wulatelong gobiensis on the basis of three “autapomorphies,” numbered below in continuation of the characters above:
11. ENF ventral margin located below mid-height of premaxilla;
12. strap-like caudal ramus (jugal process) of maxilla extends further caudally than preorbital bar (lacrimal), and overlaps the jugal;
13. rostrodorsal process of surangular constricted dorsoventrally in lateral view.
To evaluate these, I compared the presumed autapomorphic features among specimens of Oviraptoridae, focusing on the skull. Reference was also made to another new oviraptorid, Ganzhousaurus nankangensis (Wang, Sun, Sullivan & Xu, 2013), described from a specimen that had been collected by private persons and is purported to have derived from the Nankang Basin of Jiangxi Province, China. This suggests that it derives from the region of collection for the oviraptorids Heyuannia huangi and Banji long, the therizinosauroid Nanshiungosaurus brevispinus, tyrannosaurids and hadrosaurids, reflecting similar faunae and depositional environments as is found in the Nemegt Formation and Horseshoe Canyon Formation, and thus the fauna may be of latest Campanian or early Maastricthian in age. But because the provenance is unknown, and the dealer was reluctant to reveal details of its collection, doubt arises around the discovery, collection and handling of the specimen prior to acquisition by the Shandong Museum of Jinan Province, where the specimen is reposited as SDM 2009-03-02.
1 & 11. Position of the external narial fenestra (ENF). In oviraptorids, the external narial fenestra is a large, oval fenestra; it bordered by the nasal and the premaxilla around its entirely circumference, but the association of other bones depends larger on where it is in the skull. An exceptionally deep subnarial premaxillary “corpus” (or body) [also noted here] occurs in all oviraptorids, but how deep depends. In my previous post that discusses that subject, I noted that relative height is poorly quantified, and this is also the case here. Xu et al. do not show relative height of the ENF within the premaxilla or in relation to the rest of the skull, and indeed it is difficult for them to do so, as the portion of the premaxilla that forms the rostral and dorsal margins of the ENF are missing. But it is possible to use a proxy for relative height, and that is the long axis of the skull, here determined along the ventral margin of the jugal (which in virtually all oviraptorids is generally straight along its ventral margin; I discussed this in more detail here). When this line is drawn through the skull, it is clear how the naris is positioned on the skull merely by is ventral margin being preserved.
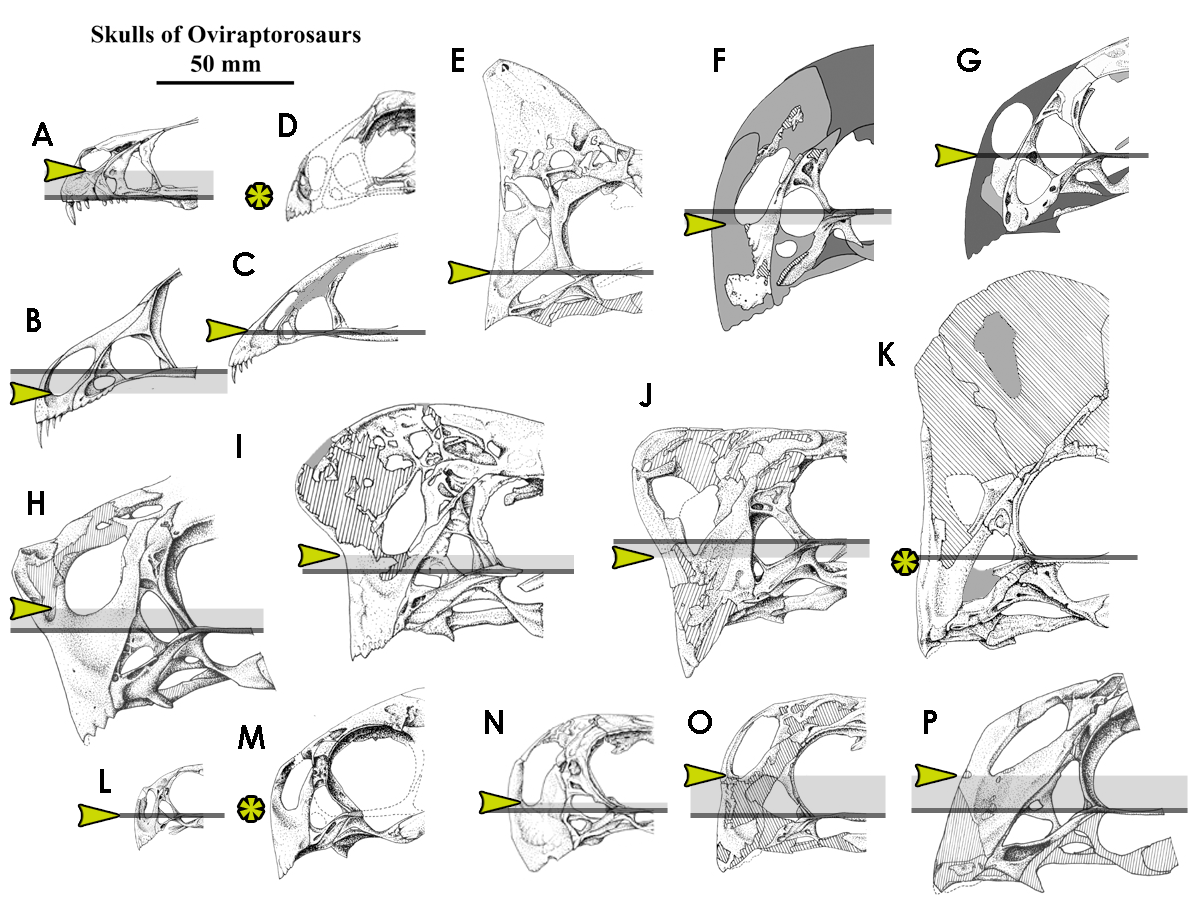
A chart of snouts for oviraptorosaurs; all illustrations produced by me. A, Incisivosaurus gauthieri; B, Caudipteryx sp.; C, Caudipteryx zoui, grey parts restored; D, Avimimus portentosus; E, “Mitrata” oviraptorid, undescribed; F, Wulatelong gobiensis; G, Oviraptor philoceratops; H, Citipati osmolskae; I, MPC-D 100/42; J, Nemegtomaia barsboldi; K, Rinchenia mongoliensis; L, Banji long; M, ZPAL MgD-I/95, often referred to “Ingenia” mongoliensis, but a conchoraptorine instead; O, Conchoraptor gracilis; P, Khaan mckennai. All skulls are to scale. Horizontal grey lines reflect the ventral jugal margin, which approximates “natural attitude” of the head at rest; orange-ish arrows point to the level of the ventral margin of the narial fenestra, and the grey box illustrates the extent to which this expands above or below the line. Asterisks indicate lack of reliable information.
Relative to an assumption of how high the premaxilla would be in IVPP V18409, I estimate that the ventral margin of the ENF is positioned at 46% of the premaxilla height. This position is 50% for a large range of oviraptorids, including Incisivosaurus gauthieri, the Zamynkhondt oviraptorine, and Nemegtomaia barsboldi; it is higher (which equates to a position higher relative to premaxilla height) in Citipati osmolskae (64%), Banji long and Khaan mckennai (both 63%), the “Big Beak” conchoraptorine and ZPAL MgD-I/95 (often referred to “Ingenia” yanshini, as it is assumed in Xu et al. (2013), which are at 57 and 56%, respectively); but is also low, as in Caudipteryx (41% for both Caudipteryx zoui and sp.), Rinchenia mongoliensis (40%), Yulong mini (43%) or the “mitrata citipatiine” (20%). As such, given that the position of the ENF varies above and below the 50% point among oviraptorids (although this is probably due to the size of the crest), it is questionable whether the value is meaningful. However, it is lower than most oviraptorids, some of which (e.g., Yulong mini) are probably basal oviraptorids[n2], along with Oviraptor philoceratops, and outside the “citipatiine” and “ingeniine” distributions. If I’ve overcalculated the height of the premaxilla, and have included instead portions of the nasal or frontal, then the ENF is much higher relative to premaxilla height; but alternatively, like Oviraptor philoceratops, Rinchenia mongoliensis and the mitrata oviraptorid, as well as the hell Creek caenagnathid, it is also possible that Wulatelong gobiensis sports a tall cranial crest.
2 & 3. Proportion of the orbit and shape and proportion of the infratemporal fenestra. The shape and size of the orbit is generally constrained among oviraptorids, where it roughly ovoid, with the long axis of the orbital fenestra aligned to the skull long axis. It is remarkably circular in Khaan meckennai and Banji long, but more ovate in most other taxa. The infratemporal fenestra, on the other hand, varies quite a bit among oviraptorids. This is due in part to the lack of a definite caudodorsal margin on the lateral surface of the squamosal, which extends a long process down the lateral surface of the quadrate rostral to the quadratojugal process, thus enclosing the proximal end of the quadrate. The ITF is broadly rectangular and almost always higher than rostrocaudally long, but this measure is made by drawing a line parallel to the jugal ventral margin axis and positioned at the rostralmost extent of the orbit. This line typically meets the contact between squamosal and quadratojugal, but this position can vary.

A, Wulatelong gobiensis; B, MPC-D 100/42; C, Citipati osmolskae; D, Yulong mini; E, “Big Beak” conchoraptorine; F, Rinchenia mongoliensis; G, Nemegtomaia barsboldi; H, Oviraptor mongoliensis. A is from Xu et al. (2013), B, C, E and F from Osmólska et al. (2004), D is from Lü et al. (2013), and H is from me. Skulls are not to scale. Proportions of the orbit and infratemporal fenestrae. Line drawn from midheight of caudal margin of infratemporal fenestra (roughly at the dorsal limit of the quadratojugal where it meets the squamosal) to the rostral end of the orbit; line is approximately even to the ventral jugal margin, which approximates “natural attitude” of the head at rest.
Measuring the segments of this line only within the ITF and orbit, an estimate of rostrocaudal length is derived, which can be compared relative to each other. At the same time, a rough estimate was made of the dorsal, rostral, ventral and caudal margins as determined by rough approximation of the largest rectangle that can fit within the proscribed ITF. Note that this rectangle is not equivalent to a frame composed of the greatest rostrocaudal “width” and dorsoventral “height,” and it measures instead the relative slopes and angles of rami that form the ITF’s margins. These are shown above as lower-case letters of their associated images, in red; g is shown in lighter color this margin is speculative, and based on broad assumptions of the missing squamosal. First, the orbit is longer than the ITF on the rostrocaudal axis in all shown taxa with the exception of Rinchenia mongoliensis; the ITF varies between 82 and 89% of the oribtal length in all taxa save Rinchenia mongoliensis (108%) and the “Big Beak” conchoraptorine, in which the ITF is nearly half (53%) of the orbit length. (Not shown is Khaan mckennai, 79%.) There is no significant difference among noted taxa as they vary within 10%, and in comparison to the full range, Wulatelong gobiensis (85%) compares best with Oviraptor philoceratops (87%), Yulong mini (82%), and Citipati osmolskae (88%). Thus, it is not likely that this value qualifies a basal-derived split. Xu et al. (2013) also noted that the dorsal border of the ITF is much shorter than the ventral border, which creates a skewed, trapezoidal box (see c, d, above). Also not shown is Khaan mckennai, which resembles c and d most. Because a very short dorsal margin relative to the ventral occurs in other oviraptorids, both derived and basal, it is also not likely this character qualifies a split amongst taxa. Both taken into account, the ITF shape and size are relatively similar to other citipatiine oviraptorids, but also conchoraptorine and Oviraptor philoceratops. A more rectangular, less trapezoidal patten, as in Nemegtomaia barsboldi (h), appears to be the exception, not the rule.
4, 5 & 12. Proportions of the lacrimal, extent of maxilla and position of ectopterygoid. Xu et al. (2013) argued that an autapomorphic feature of IVPP V18409 was the caudal extent of the maxilla relative to the lacrimal and ectopterygoid, as well as the the supposedly forward position of the ectopterygoid relative to the lacrimal. They further indicated that the proportions of the maxillary (rostral) and orbital (caudal) processes of the lacrimal suggested a basal alignment. Examining the data presented in the figure when discussing the first few characters, note the orange lines forming a “┼.” This shape describes the long, vertical axis of the lacrimal, and the crossbar formed by drawing a line from the rostralmost to the caudalmost extent of the processes. This line also passes through the vertical process of the ectopterygoid, demonstrating alignment of these bones, in most taxa.

A, Wulatelong gobiensis; B, MPC-D 100/42; C, Citipati osmolskae; D, Yulong mini; E, “Big Beak” conchoraptorine; F, Rinchenia mongoliensis; G, Nemegtomaia barsboldi; H, Oviraptor mongoliensis. A is from Xu et al. (2013), B, C, E and F from Osmólska et al. (2004), D is from Lü et al. (2013), and H is from me. Skulls are not to scale. Proportions and orientation of lacrimal processes. Proportions of the lacrimal rostral and caudal processes aligned to the long vertical axis of the lacrimal. ≈, rostral and caudal processes roughly equivalent; arrow points to longest process when they are not equal; asterisks in F and H indicate no reliable way to measure.
Relative length of the rostral and caudal processes of the lacrimal demonstrate that only Incisivosaurus gauthieri and Caudipteryx sp. appear to have processes of relative equal length; all other taxa for which these processes are available (leaving open the question of whether this is observable in MPC-D 100/20, holotype of Conchoraptor gracilis) have longer caudal processes, and are marked with a blue asterisk. In taxa for which the ectopterygoid can be observed, the lacrimal and ectopterygoid are aligned; medially, they form an interlocking brace, and the lacrimal sends a ventral process that articulates with the ectopterygoid medial to the jugal and maxilla. The alignment occurs as the C-shaped ectopterygoid central portion forms a beam that is generally parallel to that of the lacrimal, but sends a process along the ventral and medial jugal that reaches further caudally than the ascending process itself. In some taxa, this ascending bar is not aligned to the lacrimal, as in the “Mitrata” citipatiine (it is further caudally-placed than the lacrimal), Wulatelong gobiensis, and to a degree Rinchenia mongoliensis, in which the process is broad and can be said to be aligned but also extend further caudally than the lacrimal. The degree to which the ectopterygoid is “located caudally” is slight, and Xu et al. (2013) do not offer a way to quantify this; though while I find that there is a method to do so, this method does not strongly support their claim.

A, Wulatelong gobiensis; B, MPC-D 100/42; C, Citipati osmolskae; D, Yulong mini; E, “Big Beak” conchoraptorine; F, Rinchenia mongoliensis; G, Nemegtomaia barsboldi; H, Oviraptor mongoliensis. A is from Xu et al. (2013), B, C, E and F from Osmólska et al. (2004), D is from Lü et al. (2013), and H is from me. Skulls are not to scale. Alignment of maxilla and lacrimal. Down-pointing arrows are aligned to the upper axis of the ectopterygoid, and with the possible exception of Citipati osmolskae these lines pass through the main vertical axis of the lacrimal; shorter up-pointing arrows indicate the caudal-post position of the maxilla in lateral view (this position would be further rostral in medial view); 1 and 2 refer to the possible further caudal extent of the maxilla per Xu et al. (2013), but this position is effectively meaningless in the comparison to the sample.
Finally, Xu et al. (2013) suggest that the caudal extent of the maxilla is less rod-like and forms a broad plate, but which also extends further caudally than the lacrimal or ectopterygoid; this, however, is not apparent in the figures provided, and upon comparison of the lateral view to other specimens, it would appear that the IVPP V18409 does not exhibit any further extent of the maxilla caudally, nor the broadness of the caudal, jugal ramus; nor indeed does it show the jugal having an overlapping shape any less than in other oviraptorids. In no oviraptorid does the maxilla seem to extend further caudally in lateral view than does the lacrimal; the ectopterygoid forms a long contact with the jugal and the maxilla simultaneously in all oviraptorids, and this prevents the maxilla extending further caudally even medially, where the lacrimal overlaps the ectopterygoid.
13. Shape of the rostrodorsal process of the surangular. The anterodorsal process of the surangular is defined largely by extent of the external mandibular fenestra (EMF) caudally and ventrally to the surangular/dentary contact. This contact forms an elongated, V-shaped suture in all oviraptorids, with the dentary forming the apex of the “V” and pointing caudally. In oviraptorids, the shape of the rostrodorsal process is elongated and lies medial to the caudodorsal process of the dentary, and to the medial of this lies the coronoid. The EMF forms a shape that Pete Buchholz and I craftily referred to as heart-shaped: The dentary is divided into a dorsal and ventral process and the margin between these is deeply incised into the caudal margin of the dentary, and is always rounded, semi-circular; but the postdentary portion is more complex, with a margin bisected by an elongated, triangular process that projects roughly to the rostralmost extent of the EMF itself, separating the caudal margin into two “lobes”. This is evident below:

Relationships of the dentary, surangular and EMF in oviraptorosaurs. A-J, after Wang et al. (2013). Bottom two illustrations rendered after Xu et al. (2013) and Lü et al. (2012), respectively. Horizontal black line indicates the greatlest long axis of the mandible, while the cross-line refers to the axis of the caudal margin of the EMF, drawn across the caudal ends of the dorsal and ventral “lobes”. Numbers below each figure refer to relative proportion of the postdentary bones to the mandible length (left) and relative length of the EMF to the mandible length. Black arrows point to constriction of dentary/surangular “bar”, while blue arrows refer to a large distance between the caudal EMF axis and the caudodorsal process of the surangular. A constriction with spaced process are only present together in Heyuannia huangi (b) and Wulatelong gobiensis. Mandibles are not to scale.
Xu et al. (2013) argue that the dorsal process of the surangular is constricted in lateral view, and this is correct. However, it is also constricted in ZPAL MgD-I/95 (Osmólska, 1976, which is commonly referred to “Ingenia” yanshini), MPC-D 100/978 (holotype of Citipati osmolskae), MPC-D 100/32A (holotype of Rinchenia mongoliensis) and, to a degree, in CMN 8776 (holotype of Caenagnathus collinsi), and a non-oviraptorid. A further peculiarity is the distance between the caudal margin of the EMF and the caudal extent of the posterodorsal process of the dentary; these are well-separated in AMNH FABR 6517 (holotype of Oviraptor philoceratops), ZPAL MgD-I/95, HYMV1-1 (holotype of Heyuannia huangi), which itself lacks the constriction, MPC-D 100/32A and IVPP V18409 (holotype of Wulatelong gobiensis). The morphology of the region is not explicitly incomparable across a broad enough range of taxa to qualify as “autapomorphic.”
6. Size, shape and relationship of external mandibular fenestra (EMF). In reference to the image above, the EMF has this peculiar shape. I have droaw a line across the caudal extents of the EMF’s dorsal and ventral lobes; I have also taken the liberty of applying this line to Caenagnathus collinsi, where it seems the “heart-shape” may be a derived rotation of the more basal “kidney-shape” in taxa such as caenagnathids and Caudipteryx. This line helps demarcate the two possible caudal extents of the EMF, and thus the “length” of it.
I have chosen to qualify measurements using it using the longest rostrocaudal distance that passes through the ventral lobe of the EMF, parallel to the long axis of the mandible as described by the longest line that can pass through the mandible. You will note that this departs from my typical mandibular orientation scheme, because I am measuring in regards to someone else’s figure. Adjustments are easy with a reorientation, but require validation for that methodology; as such, it is not employed. Two values were produced, the first being the post-EMF length of the mandible as a proportion of the total mandibular length, and the proportion of the EMF to the total mandibular length. These values are shown at left and right below each mandible shown above. (Not shown is Rinchenia mongoliensis, where these values are 51 and 22%, respectively.) In comparison to other oviraptorids, Wulatelong gobiensis compares very well with Oviraptor philoceratops and Rinchenia mongoliensis in the post-EMF proportion of the jaw, though this is based on estimated jaw length, and somewhat more broadly with Ganzhousaurus nankangensis (Wang et al., 2012) and Khaan mckennai. It is broadly distinct from Yulong mini (43.6%), and even further from “citipatiine” oviraptorids, whose values range above 55%, showing the EMF is restricted rostrally in the jaw (this number is highest in Nemegtomaia barsboldi, whose value at 62% coincides with coossification of the articular to the surangular, also seen in Yulong mini and caenagnathids, and an implied basal trait. Even making an assumption by reconstructing the jaw in agreement with the “citipatiine” condition, the EMF seems rather caudally-positioned.
Length of the EMF relative to the jaw is not useful with regards to Wulatelong gobiensis, as the mandible preserves so little rostral to the surangular as to make reconstruction foolish (though I do offer some dotted lines based on the “citipatiine” morphology). However, basal oviraptorosaurs have fairly large EMF proportions, with Yulong mini and Oviraptor philoceratops having values above 30%, a number that is approached by other oviraptorids, but never reached (18-27%). Caenagnathus collinsi, however, has a value only at 29%, a value below the threshhold and equidistant between Khaan mckennai (27%) and Oviraptor philoceratops (31%). While 30% seems a fairly useful value to mark the transition between the longer caenagnathid morphology that appears to be basal to Caenagnathidae and Oviraptoridae, and the derived “citipatiine”/”ingeniine”/ “conchoraptorine” conditions, which shrink down to 25% or lower, it is not possible to affirm the value I estimate above for Wulatelong gobiensis at 31%. On its face, though Wulatelong gobiensis does seem to have a relatively caudally-positioned EMF, but this doesn’t seem to mean much.
7. Slender and short scapula. This character can only be compared to Citipati osmolskae, Khaan mckennai, Rinchenia mongoliensis, Nemegtomaia barsboldi, Oviraptor philoceratops, and Heyuannia huangi. The scapula is “slender,” in that the dorsal and ventral margins do not diverge distally towards the end very much along the blade’s length, though in all taxa the distal end is expanded. In Heyuannia huangi and Rinchenia mongoliensis, this morphology is similar, and the blade is relatively narrow; in other taxa, the blade margins diverge more strongly nearer the proximal end.
8 & 9. Proportion of the pubic peduncle and size of the ischium. Unfortunately, the ilium in Oviraptoridae is poorly described, owing to the tendency to leave the ilium entombed in rock rather than prepping it out (the work of Balanoff & Norell, 2013, is unique in that they fully prepared and presented the ilium of Khaan mckennai in multiple views, although to be fair it was an isolated element; the ilium of Nemegtomaia barsboldi and Avimimus portentosus are equally well-presented, but poorly visualized). While this is mostly due to the rather Swiss-cheese-esque condition the bones are in when derived from the Gobi, it makes comparison of various shapes and perspectives difficult. In the only ilia for non-oviraptorids known, the pubic peduncle is narrow, and not as broad as the ischiadic peduncle, which forms a broad antitroochanter for the femur (Currie & Russell, 1988). This morphology is based on an ilium that has been crushed, and it is not clear if the shape of the ilium in this specimen is as in life, thus lending doubt to its proportions in comparison to oviraptorids. Caudipteryx zoui and sp. demonstrate well-preserved ilia, but to my knowledge these elements have not been fully prepped and may not accurately describe their shape.
In Avimimus portentosus, the pubic peduncle and the pubis are fused when known (Kurzanov, 1983, 1985), whilst the ischiadic peduncle is free; the ischiadic peduncle formas a broad antitrochanter, but the pubic peduncle seems to be only as broad, a condition that compares well to Nomingia gobiensis (Barsbold et al., 2002) and Shixinggia oblita.
In several oviraptorids, the pubic peduncle is craniocaudally short, but mediolaterally broad, and articulates the pubis in a broad but shallow symphysis. Its precise proportions are not otherwise available to me to evaluate, but it seems in Khaan mckennai the pubic peduncle is not merely inflected medially but narrower than the ischiadic. The ischium is preserved in several oviraptorids, but its full shape is unknown in most. In “Ingenia” yanshini, this forms a broadly straight element with a prominent obturator process; in Khaan mckennai, a similar shpae is found, but with a larger obturoator process bearing a distinct, slightly hooked distal end. In “Ingenia” yanshini, the ischium is relatively long, nearly the length of the ilium (82%), whilst in Khaan mckennai it is shorter (65%). A nearly complete pair of ilia, ischia and pubes are preserved in HMV1-1, holotype of Heyuannia huangi, and show that the ischium, which agrees in shape with Khaan mckennai, are relatively longer (63% compared to 65%) and more strongly curved than those of Khaan mckennai. The ishium in Citipati osmolskae is preserved only in its distal end in one described specimen (MPC-D 100/978), and their shape in lateral view or their length are unknown; however, the position of the ischia are seen in life position in comparison to the rest of the skeleton, including the pubes and femoral, demonstrating a maximum length appears to approximate iliac length, and are thus comparable in length to “ingeniine” taxa. The ischia are unknown in Nemegtomaia barsboldi and Rinchenia mongoliensis, nor are they present in Shixinggia oblita. Ischia for non-oviraptorid are known in Nomingia gobiensis, the ischium is strongly curved, forming nearly a L-shape in lateral view and with a simple triangular obturator; the total length of the ischium is 55% that of the ilium.
In Wulatelong gobiensis, the ischium is incomplete, but what is apparent is that the distal end tapers strongly after the obturator process, and suggests the obturator’s position relative to the pubis and ischium, in reasonable articulation, would produce a fairly short ischium, but not as short as Nomingia gobiensis‘, as the ischium (being generous with the distal end) may be estimated at 65% of the iliac length, and thus agrees with Khaan mckennai proportionately. In contrast, the ischia of Caudipteryx spp. range from 73% (zoui), 85% (dongi, probably a smaller individual of zoui) and 56% (sp., which probably needs a new name by now), and they are almost always neatly L-shaped. The proportion in Similicaudipteryx yixianensis (a probable caenganathid) is 76%, and like Caudipteryx and Chirostenotes pergracilis (51%) and Epichristenotes curriei, the ischium is L-shaped.
To constrain these measurements, length of the ischium was also compared to the pubis, and then the pubis to the ischium. In these, the values for Wulatelong gobiensis generally agree with non-oviraptorid caenagnathoids, such as Nomingia gobiensis (isch:ili 59%, isc:pub 58%) and Chirostenotes pergracilis, whilst other oviraptorids have isc:pub values at 74% (Khaan mckennai) or higher (“Ingenia” yanshini at 85%, Heyuannia huangi at 83%). The pubis:ilium values also show a dichotomy, with “ingeniine” oviraptorids have values between 79% (Heyuannia huangi) and 97% (“Ingenia” yanshini), which is not that much different from Wulatelong gobiensis (108%). High pubic length occurs in Caudipteryx spp. and all other non-oviraptorids, in which it either approaches or far exceeds the length of the ilium (130% in Epichirostenotes curriei, estimating the length of the pubis in ROM 43250); in oviraptorids, this is approached, but not exceeded, although this is true also of Avimimus portentotus (pub:ili 92%, isc:pub 66%). I would conclude that the ischium length is not necessarily useful here, and that Xu et al. (2013) chose to use it based on the presumption of limited comparison, while the length of the pubis seems far more valuable to asserting a basal position for Wulatelong gobiensis. Despite this, the proportions of the ilium and ischium appear within the range of non-basal oviraptorids, and thus that the data above is ambiguous.
10. Strongly “pinched” MTIII. This one is correct; only a slight narrowing of the proximal end of MTIII occurs in most Oviraptoridae, on which basis it may be presumed that a caenagnathid/caudipterid-like subarctometatarsus is present, the first for Oviraptoridae. However, the condition in Khaan mckennai approaches that seen in Wulatelong gobiensis, and this factor may qualify this character as a gradational feature needing further qualification. As the visibility and orientation of the pes in IVPP V18409 is problematic (the pes seems twisted slightly medially), it is tricky whether using the photographs provided in the paper are useful in measuring the diameter of the pes. It seems further that MTIV is also much broader at the midshaft than are either MTIII or II, and if this is the case (a convergence of perhaps some meaning with Troodontidae), this is another potentially useful diagnostic character, but again requires a better view. It is my opinion the current photograph is not useful in clarifying this issue.
Other Factors
As with the pes, much of what seems interesting about Wulatelong gobiensis seems to be what the authors did not speak about. This includes not only the structure of the skull in specific but of the ilium, and the large vertebral pneumatopores. Based on casual examination of other oviraptorids, many of these features tend to strongly favor comparison to Rinchenia mongoliensis, and it is with this that I agree that Wulatelong gobiensis is diagnostic, but due to factors the authors did not presume: 1) presence of a subarctometatarsal pes; 2) possession of a short second toe with a pedal ungual slightly more curved than digit four (as in Citipati osmolskae); 3) manual unguals lacking proximodistal expanded “lips”; 4) manual digits 1 and 3 subequal, with manual digit 2 longer than 3 (as in “ingeniine” oviraptorids); 5) pubis exceeds ilium in length. Andrea Cau has entered Wulatelong gobiensis into his “Megamatrix,” and reports here that he found that Wulatelong is close to the base of Oviraptoridae, within a clade of basal “oviraptorines” including Citipati osmolskae, Nemegtomaia barsboldi, Rinchenia mongoliensis. It may also end up outside this group, and basal to a “inegniine”-oviraptorine division amongst oviraptorids. There will come a time when the systematics and nomenclature need to be seriously emended for Oviraptoridae, given how speciose it is getting, but that will have to wait until “Ingenia” gets a new name.
 Examining the skeleton for meaningful features, it is clear that Wulatelong gobiensis is a truly mosaic animal. Above is a chart, using the small-headed, natural skeleton as an example, of numerous characteristics that seem to pull Wulatelong gobiensis closer to caenagnathids, yet it still has oviraptorid characters, which in this case would be plesiomorphies for Oviraptoridae.
Examining the skeleton for meaningful features, it is clear that Wulatelong gobiensis is a truly mosaic animal. Above is a chart, using the small-headed, natural skeleton as an example, of numerous characteristics that seem to pull Wulatelong gobiensis closer to caenagnathids, yet it still has oviraptorid characters, which in this case would be plesiomorphies for Oviraptoridae.
These characteristics are: 1) small size of the skull; 2) deeply everted palatal bones; 3) dentary/surangular contact rounded, not S-shaped; 4) expansive, “heart-shaped” EMF; 5) posterodorsal process of dentary set apart from caudal margin of EMF, with “pinched” surangular; 6) scapula doesn’t expand strongly towards end; 7) large brachial fossa on coracoid; 8) large sternum; 9) metacarpal III much shorter than metacarpal II, equal to I; 10) nonungual phalanges of manual digit 3 relatively equal in length; 11) no lips on unguals; 12) proportions of manual digits; 13) ilium dorsal margin peaked over acetabulum, rather than level or elevated toward cranial end; 14) pubis long and relatively straight below half-length; 15) ischium much, much shorter than pubis; 16) narrow subarctometatarsal pes; 17) very broad MTIV; 18) large, hyperextensible toe?
Some of these things are pretty interesting. The ilium has a dorsally convex margin, but the “peak” is above the acetabulum, rather than the preacteabular peak seen in Chirostenotes pergracilis (RTMP 79.20.1), Nomingia gobiensis (MPC-D 100/119), or even Rinchenia mongoliensis (MPC-D 100/32A). The pubis is very long, and the ischium very short relative to the ilium, a factor that also agrees with basal oviraptorids such as “Caudipteridae” and Caenagnathidae. The head, apparently quite small, fits into a basic picture amongst oviraptorosaurs and suggests the relatively large heads of the Hell Creek caenagnathid and Oviraptor philoceratops are autapomorphically large. The hand looks like a cross between a caenagnathid and an “ingeniine” oviraptorid. The foot suggests that oviraptorids may have started out with a far more robust, narrower ankle morphology as in “elmisaurids,” but with its seeming hyperflexible toe it may have been a bit more predatory than a typical oviraptorid seems. Strange. These features are almost all now certainly symplesiomorphic of Caenagnathoidea, and Wulatelong gobiensis approaches the quintessential “first oviraptorid.”
This animal is the closest to the first oviraptorid discovered so far, and this despite living at the same time as taxa like Khaan mckennai, which is relatively derived as oviraptorids go. No oviraptorid has been recovered older than the Djadokhta-equivalent strata of Mongolia/China, but it seems likelier that some will.
[n1] For which I arbitrarily at the moment refer all other Archaeopteryx-like taxa, Wellnhoferia grandis (Solnhofen specimen), Jurapteryx recurva (Eichstätt), Archaeopteryx siemensii (Berlin, ?Thermopolis), Archaeopteryx bavarica (München).
[n2] Yulong mini is based on a series of juvenile specimens, consisting of four partial to nearly complete skulls and with some postcranial remains. Long-bone thin sections were produced (Lü et al., 2013) and none of the specimens are older than 2 years, and in comparison to taxa like Citipati osmolskae, this suggests they are very young indeed. This may influence their phylogenetic relationships, as juvenile animals tend to clade more basally. A possible caveat to this is that the embryonic oviraptorid, MPC-D 100/971 (Norell et al., 1994), comprises a skull which has apparent derived characteristics of the snout, but also has a shallow, less sigmoid curvature of the dentary. As such, the curvature of the dentary as a character is not strong evidence per se of basal relationships.
Barsbold R., Osmólska, H., Watabe M., Currie, P. J. & Tsogtbaatar K. 2000. A new oviraptorosaur (Dinosaur, Theropoda) from Mongolia: The first dinosaur with a pygostyle. Acta Palaeontologica Polonica 45 (1): 97-106. [PDF]
Fanti, F., Currie, P. J. & Badamgarav D. 2012. New specimens of Nemegtomaia from the Baruungoyot and Nemegt Formations (Late Cretaceous) of Mongolia. PLoS ONE 7 (2): e31330.
Lü, J.-c., Currie, P. J., Xu L., Zhang X.-l., Pu H.-y. & Jia S.-h. 2013. Chicken-sized oviraptorid dinosaurs from central China and their ontogenetic implications. Naturwissenschaften 100 (2): 165-175.
Norell, M. A., Clark, J. M., Dashzeveg D., Barsbold R., Chiappe, L. M., Davidson, A. R., McKenna, M. C. & Novacek, M. J. 1994. A theropod dinosaur embryo and the affinities of the Flaming Cliffs Dinosaur eggs. Science 266: 779-782.
Osmólska, H., Currrie, P. J. & Barsbold R. 2004. Oviraptorosauria. pp.165-183 in Weishampel, Dodson & Osmólska (eds.) The Dinosauria, 2nd Edition. University of California Press, Berkeley.
Wang S., Sun C.-k., Sullivan, C. & Xu X. 2013. A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of southern China. Zootaxa 3640 (2): 242-257.
Xu X., Tan Q.-w., Wang S., Sullivan, C., Hone, D. W. E., Han F.-l., Ma Q.-y., Tan L. & Xiao D. 2013. A new oviraptorid from the Upper Cretaceous of Nei Mongol, China, and its stratigraphic implications. Vertebrata PalAsiatica 51 (2): 85-101. [PDF]



Andrea Cau emailed me (privately and politely!) to let me know I messed up on his reporting of Wulatelong‘s position in his phylogeny. I’ve adjusted my post to better reflect the reporting, which was fudgy due to using Google to translate from the original Italian. D’oh. Andrea: I know you sent this privately, but I feel it necessary to have a reason for altering the text on the blog.
;-)
Pingback: Not Another One… | The Bite Stuff