It occurs to me I need a single place to collate all my various “infographic” art pieces discussing a variety of topics, so here we go. Brief summaries of their content when not obvious will follow.
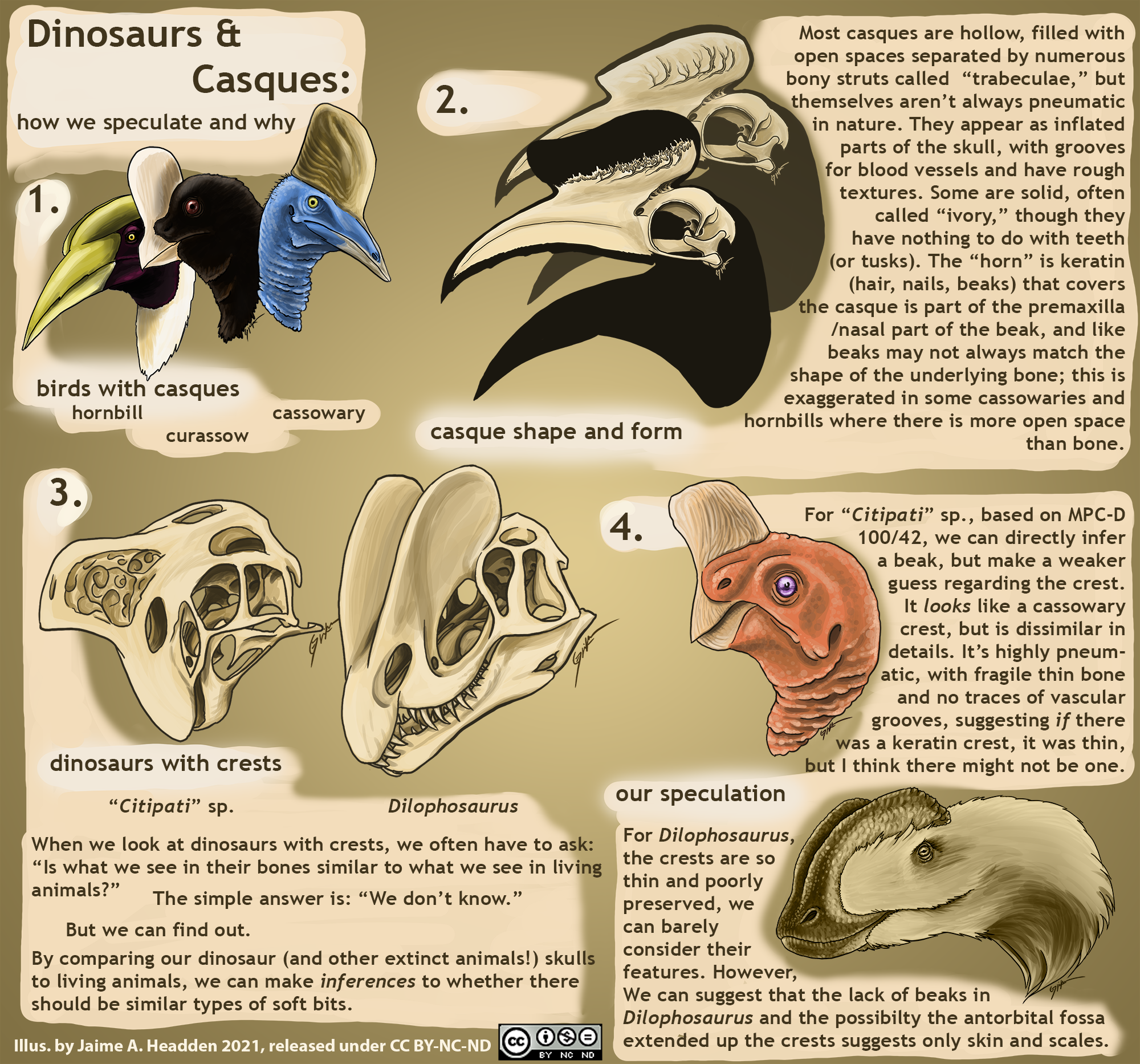

It occurs to me I need a single place to collate all my various “infographic” art pieces discussing a variety of topics, so here we go. Brief summaries of their content when not obvious will follow.
A recently completed art project (ostensibly to hone skills and objectively to explore reconstruction ideas) led me through the Ceratopsidae from its beginnings to its end. This was observed on Twitter, where I posted each full piece (at reduced size), but here I’d like to talk about the animals I depicted in a little more detail.
Continue readingOr, what is Oviraptor?
Originally described in 1924 by H. F. Osborn on the partial remains of a skeleton including the skull, neck, should, and forelimb, associated with some eggs inferred to belong to a ceratopsian the animal was predating, Oviraptor philoceratops languished in the margins of paleontological mythologica for nearly 60 years.
Continue reading
Unlike many of the Triassic marine reptiles, this particular beast was of Paleogene age, and represented such a novelty in its discovery. Originally framed as a giant crocodilian, its name, meaning “king lizard of Isis,” reflected the view of many mid-1800s giant fossil animals, being of reptilian nature regardless of reptilian characteristics. It wasn’t long after verifiable mammalian elements were identified, and what we know today comprises the longest-bodied archaeocete (“ancient whale,” a paraphyletic grade of pre-crown whales) known.
This follows in a series of reconstructions in which I depict marine tetrapods with elongated bodies, many of which had flipper-like limb modifications, small heads (proportionately) being adaptations to aquatic predation or feeding in some manner or other (there being many variationsof similar adaptations despite distinct ancestry).
A large number of Triassic marine reptiles have similar body plans: A barrel-shaped body, large flippers, short but slender neck, and moderate to small head. This bauplan is useful for many pelagic animals, although most Triassic sediments preserving these are near-shore shallow continental shelf regions.
This has led many observers, both past and present, to refer these animals into a few, small groups, even classically as few as one. Today, these groups are diverse and split amongst a radiation of sauropsids from the base (such as mesosuchids) to deep into stem-crocodiles (thalattosuchians), and a range amongst the nodes between (ichthyosaurs, plesiosaurs, choristoderes, placodonts, saurophargids, pachypleurosaurs, etc.).
Continue readingQuite some time ago, I fell in line with the authors in reconstructing the weird Atopodentatus unicus with a particularly Lovecraftian face. I won’t apologize for this reconstruction, I followed the implied anatomy the authors presented and couldn’t distinguish what, at the time, looked like a reasonable reconstruction but was not, in fact, correct.

I present now an improved reconstruction. (The feet aren’t even backwards as they’d been in my drawing, above…)
Continue readingBeelzebufo ampinga (Evans, Jones & Krause, 2008) may not be the largest extinct frog, but it was particularly large. Amongst the frogs, it’s amongst the hyperossified members of Ceratophryidae (Hyloidea, Anura) from the Late Cretaceous of Madagascar. There’s not much here for me to say.

Yep, that’s it. Unpreserved elements of the skeleton are rendered after Ceratophrys and similar hyloidean frogs.
Continue readingIf you wondered why I love some peculiar Triassic tetrapods like Longisquama or Sharovipteryx, it’s probably due to the high diversity and degree of questions you can ask about them, and some even provide curious answers.
So here’s the answer to the riddle above:

I mean, look at that. The megalancosaurid drepanosaur Megalancosaurus preonensis is one helluva animal.
Continue reading
I might be a little safe with this reconstruction, but it represents a further attempt to clean up and add to the number of controversial archosauromorphs for which few skeletals exist, or whose existence is owed to controversial reconstructors or miseducators seeking personal fame over accuracy.
Continue reading
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. It, and all of its files and contents -- unless otherwise noted -- are protected by this license.
License established 9/9/2012, while previously I was using a CC-BY-ND license instead.
Support this blog:

I have two websites, my DeviantArt and RedBubble accounts, which allow downloads and purchase of the imagery presented on this site. As I am a starving artist, I would appreciate support this way.
